8.1. Разнообразие восточноевропейского генофонда
|
§1. Оценки по классическим и по ДНК маркёрам: Надежная панель генов - Идентичная панель популяций - Две стороны медали — HS внутри популяций, а GST между ними — География HS по классическим и по ДНК маркёрам похожа - Хотя сами HS отличаются в два раза! - По ДНК маркёрам и генам, кодирующим белки, GST между популяциями одинаковы! — И не зависят от различий внутри популяций
§2. Сравнение с регионами Евразии: Велика ли дифференциация Восточной Европы? — «Супер» регионы и просто регионы Евразии — С запада на восток Евразии дифференциация популяций нарастает — Между Западной Европой и Сибирью — Русский генофонд вбирает большую часть изменчивости Восточной Европы - Генетика послушна географии Анализ восточноевропейского генофонда начнём с выявления размаха его изменчивости. Мы оценим обе составляющие разнообразия генофонда: и генетические различия между всеми популяциями Восточной Европы (GST), и различия индивидов внутри этих популяций (гетерозиготность HS). Такие оценки можно получить только по данным генетики. Надёжность и устойчивость этих оценок мы выясним, сопоставив показания двух равноправных свидетелей. Для этого мы получим оценки разнообразия и по ДНК, и по классическим маркёрам, а потом сравним, насколько они совпадут. Наконец, чтобы понять, велико ли это разнообразие, надо сопоставить размах различий между популяциями Восточной Европы (GST) с аналогичными оценками для соседних регионов. Круг этих вопросов и обсуждается в данном разделе. Восточноевропейские народы хорошо изучены в отношении полиморфизма целого ряда белковых и ДНК маркёров [Генофонд и геногеография ..., 2000; Лимборская и др., 2002]. Но мы уже не раз говорили о том, что по отдельному гену сложно судить о закономерностях генофонда в целом. Картины изменчивости отдельных маркёров могут резко различаться по каким-либо случайным причинам, из-за различий в их функции, из-за различий в эволюционной траектории маркёров разных типов. Словом, разные факторы микроэволюции могут быть в разной степени отражены в пространственной изменчивости разных генов. Поэтому надо от анализа отдельных генов перейти к анализу их совокупности. Чтобы в изменчивости генов обнаружить общую архитектонику генофонда, следует изучить большую панель генов. Она должна, во-первых, включить значительное число маркёров, изученных во многих популяциях; а, во-вторых, эти маркёры должны различаться в биохимическом и физиологическом отношении (детально все эти вопросы рассмотрены в Приложении). Когда такие данные - о многих разнообразных генах во многих популяциях - собраны, они анализируются методами многомерной статистики, вычленяя общие для всех генов закономерности наиболее эффективным, объективным и математически корректным способом. Итак, преимущество обобщённого анализа (одновременно, совместно по всем генам) состоит в выявлении наиболее общих, объективных закономерностей генофонда. Значимость анализа отдельных генов заключается в ином - в создании объёмной картины конкретных результатов: большого количества показателей, величин, частных закономерностей. В предыдущей части книги эти два вида анализа дополняли друг друга, позволяя видеть и «лес», и «деревья». Но для цели сравнения русского генофонда и восточноевропейского нам достаточно сравнить их «обобщённые» портреты. Поэтому мы ограничимся анализом общего уровня разнообразия, в среднем по всем генам, и лишь для ДНК маркёров вскользь затронем «индивидуальные» уровни разнообразия каждого из локусов. §1. Оценки по классическим и по ДНК маркёрамВ этом параграфе мы рассмотрим две стороны одной медали: различия индивидов внутри популяции (гетерозиготность) и различия между популяциями Восточной Европы. И оценим эти различия как по классическим, так и по ДНК маркёрам. Едва ли не самым важным для нас будет сравнение показаний этих двух «очевидцев»: оно позволит отделить устойчивые закономерности от эфемерных. Чтобы провести это сравнение корректно и получить достоверные оценки изменчивости генофонда, нам придётся часто совершать краткие экскурсы в методические вопросы - особенно расчёта межпопуляционной изменчивости. До конца этого раздела мы постараемся бегло показать читателю некоторые важнейшие уголки статистической кухни геногеографа. Тем же, кто интересуется больше результатами, чем методами, мы рекомендуем пропустить пояснения, данные мелким шрифтом. ЧТО ТАКОЕ ДИФФЕРЕНЦИАЦИЯ GST? Величина генетических различий между популяциями (то есть дифференциация генофонда, уровень межпопуляционных различий по частотам генов) обычно оценивается через величину GST. Эта степень дифференциации зависит от соотношения двух факторов микроэволюции - дрейфа генов и миграций: дрейф генов увеличивает различия популяций друг от друга, а миграции, очевидно, сближают генофонды популяций региона (см. Приложение). Поэтому уровень GST свидетельствует о балансе миграций и дрейфа генов, об интенсивности миграционных и стохастических процессов в генофонде. Малые величины GST (популяции региона генетически похожи друг на друга) обычно говорят о мощном давлении миграций и слабом дрейфе генов, а большие величины GST (популяции региона резко различаются) свидетельствуют о преобладании дрейфа генов над последствиями незначительного генного потока между популяциями. Средняя по всем изученным генам величина GST оценивает селективно-нейтральную изменчивость, избавляясь от последствий отбора, смещающих оценки GST(i) по отдельным i-тым генам (см. Приложение). Поэтому именно средняя по всем генам GST - статистика (наряду с аналогичной FST-статистикой) служит для измерения генетических различий между популяциями [Nei, 1975; Алтухов и др., 1997, Bosch et al., 2000]. Второй показатель - уровень внутрипопуляционного разнообразия (HS) - оценивает среднюю ожидаемую гетерозиготность популяций [Nei, 1975; Алтухов, 1989]. РЕПРЕЗЕНТАТИВНЫЙ НАБОР ГЕНОВ Сравнение изменчивости ДНК и классических маркёров - важнейшая задача! Постараемся решить её вдумчиво и строго. Первое требование - анализ репрезентативной панели генов, которая покажет нам истинное лицо генофонда. Оно в нашем случае выполняется, поскольку в панелях представлены самые разные типы классических и ДНК маркёров. Почему мы так считаем? Дело в том, что теоретически величина различий между популяциями не должна зависеть от панели генов, по которой анализируется генофонд. Но для этого панель генов должна быть репрезентативной - отражать свойства основной части генома, а не его своеобразных частей. Поэтому здесь мы не будем касаться однородительских маркёров - у них свой путь микроэволюции, зависящий и от много более мощного дрейфа генов, и от различий между паттерном «мужских» или «женских» миграций. А включим в анализ только аутосомные маркёры. Панель ДНК маркёров сформирована так. что включает три разных класса маркёров - инсерционно-делеционного, минисателлитного и микросателлитного полиморфизма. Это позволяет надеяться, что мы сможем избежать смещений оценок изменчивости, связанных с особенностями микроэволюции тех или иных классов маркёров (см. Приложение). Анализ ДНК маркёров проведён по 114 аллелям шести наиболее подробно изученных аутосомных локусов (табл. 8.1.1.). По классическим маркёрам число аллелей примерно такое же - 100 аллелей. А число локусов классических маркёров велико - тридцать три и среди них также есть представители трёх классов: биохимические, иммунологические и физиологические маркёры. Поэтому мы можем надеяться, что обе панели маркёров - классических и ДНК - репрезентативны. ИДЕНТИЧНЫЙ НАБОР ПОПУЛЯЦИИ Второе требование - анализ одних и тех же популяций. Его мы также выполнили, сведя массив данных о классических маркёрах к тем же народам, которые изучены по ДНК. Если, например, по ДНК маркёрам будут изучены саамы и ненцы Восточной Европы, а по классическим маркёрам народы Дагестана, то ясно, что ни о каких сравнениях не может быть и речи. Хотя и те, и другие народы относятся к населению Восточной Европы, но они не могут дать представление обо всём генофонде региона. Если мы хотим сравнить изменчивость по ДНК и по классическим маркёрам, это сравнение надо провести строго по одному и тому же кругу популяций. По ДНК маркёрам изучен меньший набор популяций, поэтому будем ориентироваться на него. Каждый ДНК маркёр немного отличается от другого по набору популяций, но в среднем изучены 28 популяций 15 народов Восточной Европы: белорусы, молдаване, русские, украинцы, коми, мари, мордва, удмурты, адыги, аварцы, башкиры, ногайцы, татары, чуваши, калмыки. Эти народы охватывают популяции от Белого моря до Чёрного и относятся к четырём лингвистическим семьям (индоевропейской, уральской, алтайской, северокавказской). Так что можно надеяться, что они дадут представление о генофонде Восточной Европы в целом. По классическим маркёрам изучен, конечно же, намного более широкий круг народов Восточной Европы. Но чтобы максимально корректно сравнить их с ДНК маркёрами, при расчётах статистических показателей (HS и GST) сузим этот круг до тех же 15 народов, что и по ДНК полиморфизму (данные по прочим народам мы не будем учитывать в анализе). Но, исключив некоторые народы, не занизим ли мы при этом уровень изменчивости? Мы проверили показатели до и после сокращения выборки народов - они оказались очень близки. Это означает, что вряд ли мы исказили общие закономерности в изменчивости генофонда Восточной Европы, ограничив рассмотрение именно этими 15 народами2. ГЕТЕРОЗИГОТНОСТЬ (HS) Убедившись в ожидаемой репрезентативности наборов маркёров и в полной сопоставимости наборов популяций, рассмотрим гетерозиготность и межпопуляционную изменчивость, рассчитанную по этим данным. Начнём с гетерозиготности. КАРТЫ HS. Генетический ландшафт гетерозиготности ДНК маркёров в ареале Восточной Европы показан на рис. 8.1.1. Он варьирует незначительно (0.66<HS<0.72), но закономерно. Карта демонстрирует некоторое снижение гетерозиготности к востоку (у башкир) и к югу (у адыгов), тогда как более высокие значения гетерозиготности обнаруживаются в Поволжье, а также (HS>0.69) у части русских популяций, украинцев, белорусов. Видна закономерность снижения гетерозиготности к востоку и к северу от Поволжья. Такая картина - максимум гетерозиготности у восточных славян, снижение гетерозиготности при движении на восток к Уралу и дальнейшее снижение в Сибири - хорошо известна, она показана при анализе гетерозиготности классических маркёров в Северной Евразии [Балановская, Рычков, 1997]. Построенный нами новый вариант карты гетерозиготности классических маркёров в Восточной Европе приведен на рис. 8.1.2. Можно видеть, что уровень гетерозиготности максимален на Украине, и снижается к северу и востоку. Постепенность тренда лишь отчасти нарушается в Заволжье (в районе Ветлуги и Вычегды), где гетерозиготность ниже, чем должно бы быть при идеально ровном тренде. Второе локальное нарушение заметно на Западном Кавказе. Но общий тренд гетерозиготности классических маркёров удивительно закономерен и плавен - снижение от юго-западного угла карты к северо-востоку. 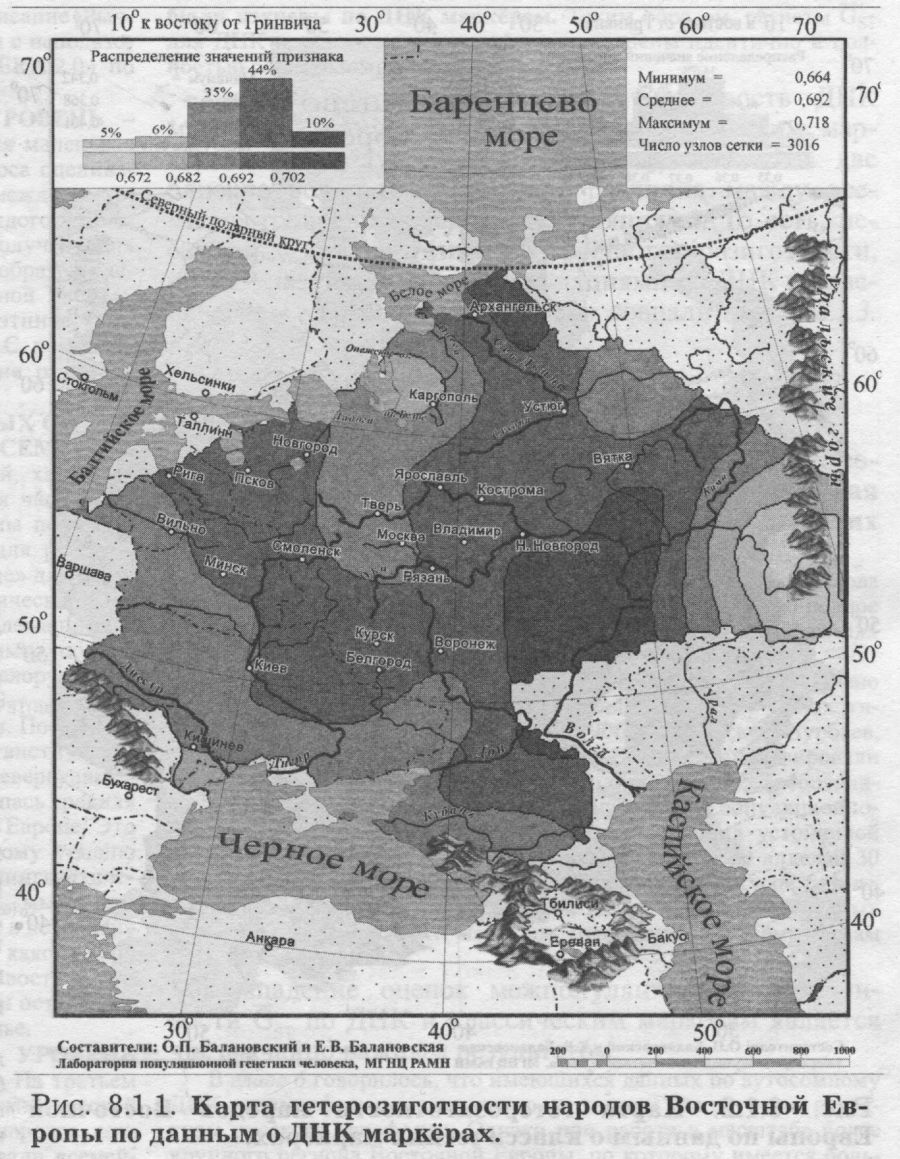
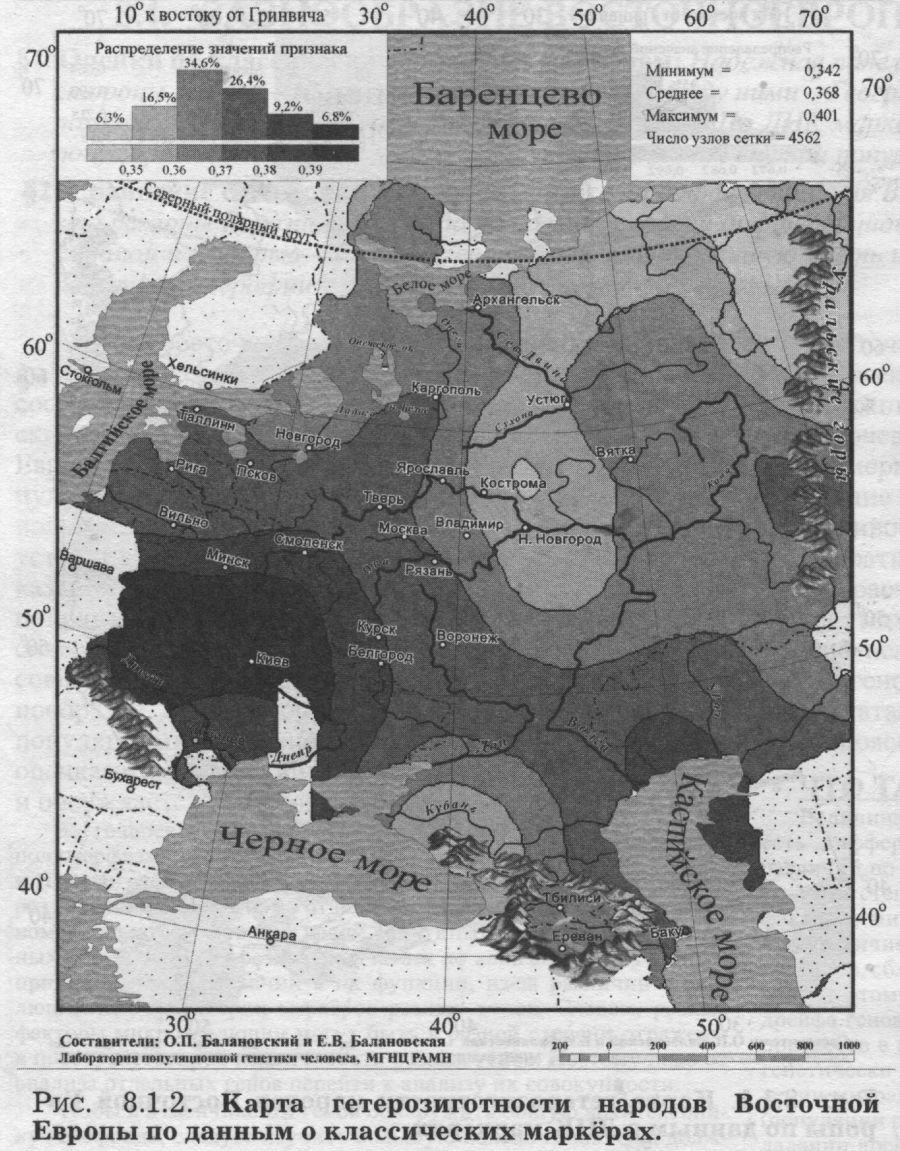
Карты гетерозиготности ДНК и классических маркёров не слишком похожи, хотя обе показывают снижение гетерозиготности к востоку и к северу. Глядя на плавный тренд «классической» гетерозиготности, основанный на тридцати трёх локусах, и путаные извивы карты ДНК полиморфизма (всего по шести локусам), можно предположить, что именно недостаток ДНК данных приводит к различиям между картами. Это будет легко проверить, когда накопятся новые данные по аутосомному ДНК полиморфизму. 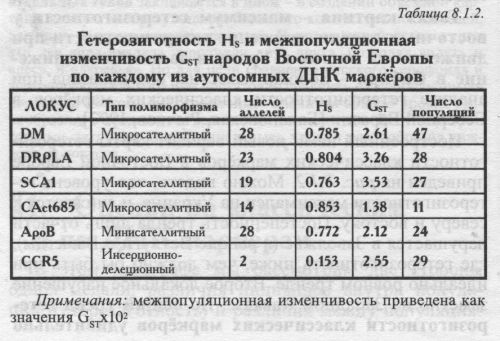
Попутно отметим, что в отличие от гетерозиготности, оценки межпопуляционной изменчивости GST отдельных ДНК маркёров .никак не связаны с типом полиморфизма. Так, и локус с наибольшей величиной GST и локус с наименьшей GST относятся к микросателлитным. СРЕДНИЕ ВЕЛИЧИНЫ HS. Но что удивительно - при общем сходстве в географии, сам уровень гетерозиготности по ДНК и по классическим маркёрам различается в два раза! По ДНК маркёрам он очень велик: HS=0.69. По классическим маркёрам оценки в два раза ниже: HS=0.34 - если считать по всем изученным народам Восточной Европы; и HS=0.37 - по тем 15 народам, которые изучены по ДНК. Конечно же, высокая гетерозиготность ДНК маркёров вызвана мультиаллельностью большинства использованных локусов (микро- и минисателлитных): из таблицы 8.1.1. видно, что классические маркёры в среднем изучены по 3 аллелям локуса, а ДНК маркёры - каждый локус в среднем по 19 аллелям. Таблица 8.1.2. показывает связь гетерозиготности с типом маркёра и с числом аллелей - у мультиаллельных локусов (микро- и минисателлитных) гетерозиготность намного выше, чем у диаллельных (их гетерозиготность не может превышать HS=0.5). Итак, при сравнении классических и ДНК маркёров нами обнаружена независимость географии (паттерна) гетерозиготности от её величины, от размаха разнообразия: величина гетерозиготности ДНК и классических маркёров различается в два раза, а изменчивость этой величины в пространстве следует сходным закономерностям. В современной геногеографии это уже не удивляет исследователя. Столь же большое сходство было выявлено нами между картами изменчивости генофонда мари по генетическим маркёрам и по генам наследственной патологии [Балановский и др., 1999; Наследственные болезни в популяциях человека, 2002], в то время как уровень изменчивости этих систем генов различался не в два раза, а на два порядке! РАЗЛИЧИЯ МЕЖДУ ПОПУЛЯЦИЯМИ GST Впрочем, гетерозиготность HS служит лишь вспомогательным параметром, оценивающим изменчивость индивидов внутри популяций. Главным же инструментом популяционно-генетического анализа выступают различия между популяциями (GST). GST отвечает на вопрос: насколько разные популяции Восточной Европы отличаются друг от друга? Чтобы рассчитать GST максимально корректно, мы учли основные популяционные «матрёшки» (см. главу 5): популяция - этнос - языковая семья - регион. Это означает, что мы учли иерархию популяций в регионе и избежали смешения оценок в сторону подробно изученных народов и целых лингвистических семей (см. Приложение). Ведь народов, говорящих на языках индоевропейской семьи, намного больше, чем остальных, и изучены они зачастую подробнее. Поэтому - без иерархического анализа, без учёта «матрёшек» мы сместили бы все значения средних частот и GST в «индоевропейскую» сторону. А это свело бы всю историю Европы к истории одних лишь индоевропейцев! КАК ПРАВИЛЬНО РАССЧИТАТЬ GST Хотя иерархический вариант расчёта GST является основным и наиболее строгим [Nei, 1975], из-за его сложности он применяется относительно редко. Поэтому приведём его краткое описание [Рынков. Ящук (Балановская), 1980]. Весь анализ проведён с использованием оригинального программного пакета «MEGERA 2.0» по следующему алгоритму. ПЕРВАЯ «МАТРЁШКА»: ЭТНИЧЕСКИЙ УРОВЕНЬ - ПОПУЛЯЦИИ В ПРЕДЕЛАХ ЭТНОСА. Это самая маленькая «матрёшка». На этом первом этапе для каждого этноса оценивалась средняя по всем аллелям величина различий между всеми популяциями данного народа, т. е. гетерогенность каждого этноса: GST(popul-ethnic) - После того как такие величины были получены для каждого народа, эти величины усреднялись, и таким образом рассчитывалась средняя гетерогенность этноса Восточной Европы. Это значение GST(popul-ethnic) присваивалось нижнему, этническому уровню иерархической популяционной структуры. С помощью этой величины GST(popul-ethnic) мы оцениваем средние различия между популяциями в «среднем» этносе. ВТОРАЯ «МАТРЁШКА»: УРОВЕНЬ ЯЗЫКОВЫХ СЕМЕЙ - ЭТНОСЫ В ПРЕДЕЛАХ ЛИНВИСТИЧЕСКОЙ СЕМЬИ. На втором этапе вычислялись средние частоты аллелей, характерные для каждого народа (как невзвешенная средняя частот аллеля в популяциях данного народа). В результате мы получали «этнические» частоты аллелей - «среднерусские» для русских, «среднеукраинские» для украинцев, «среднемарийские» для мари, и так для каждого народа. На основании таких «этнических» частот оценивались различия между народами, принадлежащими к данной лингвистической семье: GST(ethnic-lingvo) - Например, по различиям между средними русскими, украинскими, белорусскими и молдавскими частотами оценивалась величина GST(lingvo) Для восточноевропейских народов индоевропейской семьи. После того как такие величины были получены для каждой лингвистической семьи (индоевропейской, уральской, алтайской и северокавказской), усреднением этих четырех величин рассчитывалась средняя гетерогенность лингвистической семьи в Восточной Европе. Это среднее значение GST(ethnic-lingvo) присваивалось второму уровню иерархической популяционной структуры: уровню лингвистических семей. С помощью этой величины GST(ethnic-lingvo) мы оцениваем средние генетические различия между этносами в «средней» лингвистической семье. В случаях, когда данные для какого-либо народа по данному локусу отсутствовали, то изменчивость в пределах лингвистической семьи рассчитывалась по всем остальным народам, относящимся к данной лингвистической семье. ТРЕТЬЯ «МАТРЁШКА»: РЕГИОНАЛЬНЫЙ УРОВЕНЬ - ЯЗЫКОВЫЕ СЕМЬИ В ПРЕДЕЛАХ РЕГИОНА. На третьем этапе вычислялись частоты аллелей для каждой лингвистической семьи (как невзвешенная средняя частот аллеля у народов данной лингвистической семьи). В результате мы получали «семейные» частоты аллелей - «индоевропейские» для индоевропейской лингвистической семьи, «уральские» для уральской языковой семьи, «алтайские» для алтайской семьи, «северокавказскис» для северокавказской семьи. Например, усреднением «среднерусских», «среднеукраинских», «среднебелорусских» и «средне-молдаванских» частот (полученных на прошлом этапе расчёта) получена «среднеиндоевропейская» частота. На основании таких «семейных» частот рассчитывалась GST(lingvo-region) среди всех лингвистических семей. Это значение присваивалось высшему уровню иерархической популяционной структуры. С помощью GST(lingvo-region) величины мы оцениваем различия между лингвистическими семьями в восточноевропейском регионе. ВСЕ ТРИ МАТРЁШКИ ВМЕСТЕ: ИЗМЕНЧИВОСТЬ ПОПУЛЯЦИЙ В ПРЕДЕЛАХ РЕГИОНА. Эта «матрёшка» вмешает в себя все предыдущие, на заключительном этапе три величины GST - низшего уровня GST(popul-ethnic), среднего уровня GST(ethnic-lingvo) высшего уровня GST(lingvo-region) - суммировались благодаря аддитивным свойствам GST. Полученное значение GST(popul-region) и является искомой величиной GST, рассчитанной иерархическим методом. С помощью величины GST(popul-region) мы максимально корректно оцениваем средние различия популяций в восточноевропейском регионе. Например, благодаря иерархической процедуре множество изученных русских популяций не заслоняет малые этносы - все народы и лингвистические семьи представлены равноправно. РАСЧЕТ ДЛЯ КЛАССИЧЕСКИХ МАРКЕРОВ. С помощью точно такой же иерархической процедуры была рассчитана величина GST по классическим маркёрам. Ещё раз напомним, что в расчёт включены не все изученные по классическим маркёрам популяции Восточной Европы, а только те 15 народов, которые были изучены по ДНК маркёрам. Таким образом, расчеты GST для ДНК и классических маркёров выполнены идентично и полностью сопоставимы. Средняя межпопуляционная изменчивость ДНК маркёров составила GST=2.58. ПО классическим маркёрам мы получили величины GST=2.69. Эти две близкие величины выражают различия между всеми восточноевропейскими популяциями. То есть, несмотря на резкие отличия в уровне гетерозиготности, оценки различий между популяциями по ДНК и классическим маркёрам практически совпали (табл. 8.1.3. и рис. 8.1.3). СОВПАДЕНИЕ GST ПО ДНК И КЛАССИЧЕСКИМ МАРКЕРАМ Совпадение корректно полученных оценок GST позволяет утверждать, что средняя межпопуляционная изменчивость ДНК маркёров и генов, кодирующих белки, одинакова. Такое совпадение было предсказано в работах Ю. Г. Рычкова и Е. В. Балановской [Рычков, Балановская, 19906] и нашло полное подтверждение в многочисленных последующих исследованиях (см. например [Bowcock, Cavalli-Sforza, 1991; Bowcock et al., 1991 и др.]). Однако подобное равенство оценок может быть выявлено лишь при использовании представительной выборки обоих типов маркёров [Рычков, Балановская, 19906; Балановская, Нурбаев. 1997]. Напомним, что ещё до начала анализа мы проанализировали наши наборы маркёров и выяснили, что они отвечают требованиям репрезентативности. Начиная с классических популяционно-генетических работ, считается, что для получения устойчивой оценки различий между популяциями достаточно 50 аллелей 30 локусов. Наши же панели шире - 100 аллелей 33 локусов разных классических маркёров и 114 аллелей шести наиболее изученных в Восточной Европе ДНК маркёров, относящиеся к трём классам ДНК полиморфизма. Совладение оценок межпопуляционной изменчивости GST по ДНК и классическим маркёрам является чрезвычайно важным результатом. В главе 6 говорилось, что имеющихся данных по аутосомному ДНК полиморфизму, к сожалению, не хватает для изучения структуры русского генофонда. Однако при работе в масштабе более крупного региона Восточной Европы, по которому имеется больше данных, их оказывается достаточно для корректного анализа. И лучшее доказательство этому - совпадение величины межпопуляционной изменчивости по ДНК данным с аналогичной величиной по классическим маркёрам. Дело в том, что классические маркёры, изучавшиеся с середины XX века, исследованы весьма подробно и поэтому дают объективную оценку GST при оценке дифференциации генофонда Восточной Европы [Рычков, Балановская, 1990а,б; Балановская, Нурбаев, 1997; Генофонд и геногеография народонаселения, 2000]. Изученность ДНК маркеров много меньшая, отчего достоверность оценок GST по ДНК маркёрам требует специального обоснования. 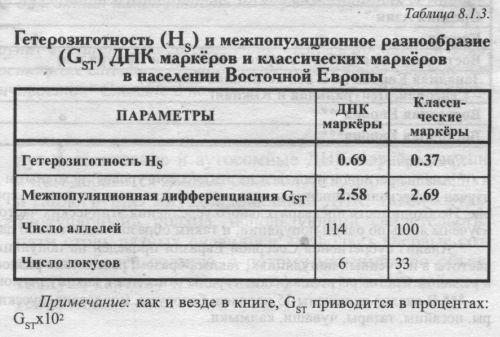
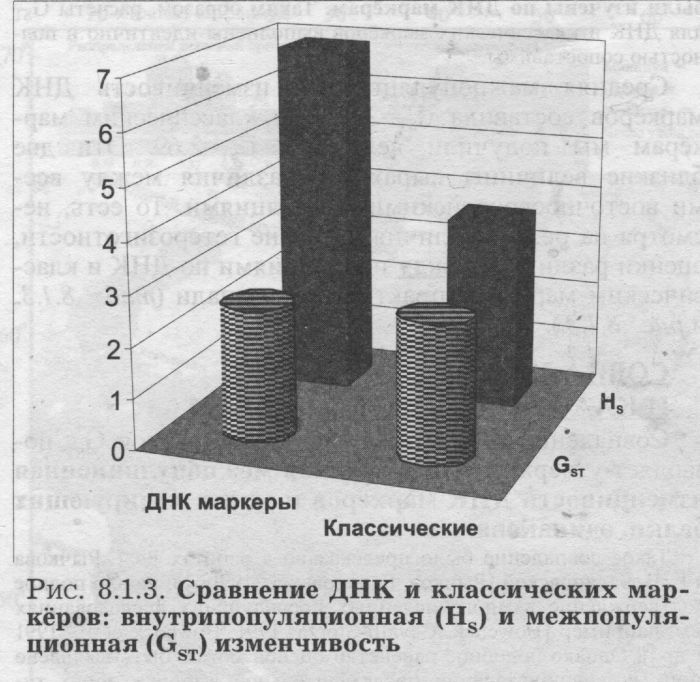
Полученное совпадение оценок GST как раз и представляет весомое доказательство, что анализируемый объем информации по ДНК маркёрам достаточен для корректной оценки дифференциации генофонда Восточной Европы. Оно указывает, что совокупность использованных ДНК маркёров даёт вполне объективную информацию о наиболее общих параметрах структуры генофонда народов Восточной Европы. И второй немаловажный вывод: классические маркёры дают надежный прогноз того, какую величину GST мы получим для аутосомных ДНК маркёров. Этот вывод мы используем уже в следующем параграфе. Выше говорилось, что сравнения межпопуляционной изменчивости ДНК и классических маркёров проводились во многих работах. В разделе 8.2. приведены результаты не только статистического, но и картографического сравнения, выполненного впервые, и вновь показавшего сходство изменчивости ДНК и классических маркёров. §2. Сравнение с регионами ЕвразииВелики или малы генетические различия между популяциями Восточной Европы? Полученная величина GST равная 2.6 или 2.7 - много это или мало? Чтобы ответить на этот вопрос, сравним её с другими регионами. «СУПЕР» РЕГИОНЫ И ПРОСТО РЕГИОНЫ ЕВРАЗИИ В таблице 8.1.4. приведены рассчитанные нами оценки межпопуляционной изменчивости GST в крупных регионах мира и субрегионах Северной Евразии (см. главу 5 и Приложение). На этом фоне удобно оценить относительный масштаб разнообразия Восточной Европы. Эти оценки межпопуляционной изменчивости регионов рассчитаны по классическим маркёрам. Благодаря совпадению оценок GST (§1) их можно использовать как прогноз для ДНК маркёров. Сначала рассмотрим самые большие «матрёшки». В таблице 8.1.4. приведены различия между народами для четырех таких «супер» регионов Евразии, масштаб которых можно оценить как субконтинентальный. Мы видим, что межэтнические оценки GST варьируют от 2.4 («супер» регион Европы и Юго-Западной Азии) до 6.8 («супер» регион Северной Евразии). 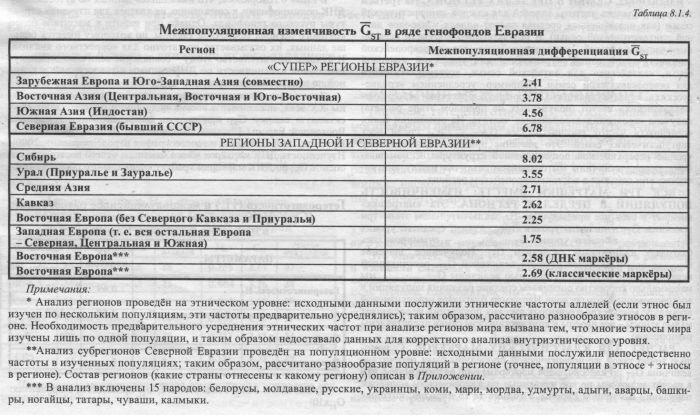
Но сейчас для нас важнее уровень пониже - «простых» регионов. Ведь эти на ранг меньшие «матрёшки» уже сопоставимы по размеру с Восточной Европой. Здесь мы учли и изменчивость популяций внутри народов (к различиям этносов внутри регионов добавились различия между популяциями внутри этносов). Сравнивая регионы Евразии, мы видим любопытную закономерность: наименьшая изменчивость характерна для населения Западной Европы (1.7), далее следуют близкие по значениям генофонды Восточной Европы, Кавказа, Средней Азии и Урала (2.2< GST <3.5); и, наибольшая изменчивость характерна для коренных народов Сибири (8.0). То есть по мере движения с крайнего запада Евразии к её крайнему востоку межпопуляционные различия нарастают. Из этого может следовать, что промежуточное положение Восточной Европы в величине изменчивости просто соответствует её промежуточному географическому положению - между Западной Европой и Сибирью. Известно, что население Европы (обычно подразумевается Западная Европа, генетически более изученная) в генетическом отношении однородно. Это положение, известное по классическим маркёрам, получило яркие подтверждения при анализе разнообразия митохондриальной ДНК. Напротив, коренное население большинства крупных регионов Азии - Южной. Восточной и особенно Северной Азии (Сибирь и Дальний Восток), характеризуются большим генетическим разнообразием популяций. ИТОГИ Для этой книги важны четыре момента, характеризующие разнообразие генофонда Восточной Европы. Во-первых, мы получили корректно сопоставимые оценки межпопуляционной изменчивости для Восточной Европы по двум типам маркёров: по классическим GST=2.7, по ДНК GST=2.6. Во-вторых, большое теоретическое значение имеет совпадение межпопуляционной изменчивости GST по ДНК и по классическим маркёрам, несмотря на их резкие различия по гетерозиготности HS. Это подтверждает возможность прогноза изменчивости ДНК маркёров по классическим маркёрам (см. Приложение). В-третьих, мы теперь знаем о промежуточном положении уровня межпопуляционной изменчивости GST Восточной Европы между уровнем Западной Европы и уровнем Сибири. Если разные популяции западной Европы генетически похожи друг на друга, то популяции Восточной Европы в полтора раза разнообразнее, а в Сибири разные популяции ещё в три раза больше отличаются друг от друга. В-четвёртых, сравнивая разнообразие восточноевропейского генофонда (GST=2.7) и русского генофонда (GST=2.0, см. раздел 5.3.) мы видим, что русский генофонд вбирает в себя значительную часть общего разнообразия всей Восточной Европы. Другие восточноевропейские народы вбирают несколько меньшую часть общего разнообразия (в среднем GST=1,2). Можно отметить, что заметная величина разнообразия русских популяций, лишь немногим уступающая разнообразию всех популяций Восточной Европы, хорошо согласуется с величиной «исконного» русского ареала, занимающего большую часть Восточной Европы. 2Такое сокращение «классических» данных до 15 народов мы. конечно же. провели только для статистического анализа - карты классических маркёров построены по полному набору имеющихся данных. |
загрузка...